��֪ʶ���ӡ������� | ��������Ӧ�����ϳ��������������

���ϳ滯ʯ
��֩����eeook.com��������֪ʶ���ӣ��� ���������߰���7��25�գ��й���ѧԺԺʿ��������ѧ������¸���֪ʶ���ӡ�Ͷ�壬��7��8��Science��־������һ���о����5.18����ijν�����Ⱥ���ϳ�Ĺ���������˲�ͬ���������� | ��¸ɣ����ȳ������ϳ棬�����Dz��Ǽ���������Դˣ������ĵ�����ͨѶ���ߡ��Ͼ���ѧ�����ѧ�빤��ѧԺ���ڽ�����������壬���������˻�Ӧ��
��֪ʶ���ӡ���7��25�շ����ˡ�����|��¸ɣ����ȳ������ϳ棬�����Dz��Ǽ������һ�ġ����У�������ѧ��¸�Ժʿ������2022��7��8�շ�����Science������[1]�����¼�� �����ġ������������������¼�� ���������������ڣ����Ҵ����о��ŶӶ� �������� �е�Ҫ������Ӧ��
���ȣ���л �������� �� �����ġ� �����˿϶��������������� ��ָ�������������������ڸ���ý�峴�÷з������ѧ�������������Ե�����һЩý��ת�����ǹ������ţ�һЩý��ת�ؾ�������Ż�Scienceͬ�ڷ������������£���������һЩ�ɷù����ǵ������У�����ʹ����һЩ�����Ͻ��ı��﷽ʽ���Դ����DZ���Ǹ�⡣ͬʱ���������� Ҳ������ ���۽ṹ�Ķ���ԡ� ����֤�����ԡ� �� ��֤�ݵĿ�ѧ�ԡ� �������������������ɡ����� �������� ��û�ж� ���۽ṹ�Ķ���ԡ� չ�����ۣ���������ͺ��������������Щ˵��������Ժʿ���������ۡ�
һЩ��������
���� �����ġ� �ĺ�������֮һ���������ϳ����������ϵ�ϵͳ����λ�ã���˵��֮ǰ�����ȼ�������ϵͳ������ϵ�������������漰��һЩ����������ڴ�����⡣
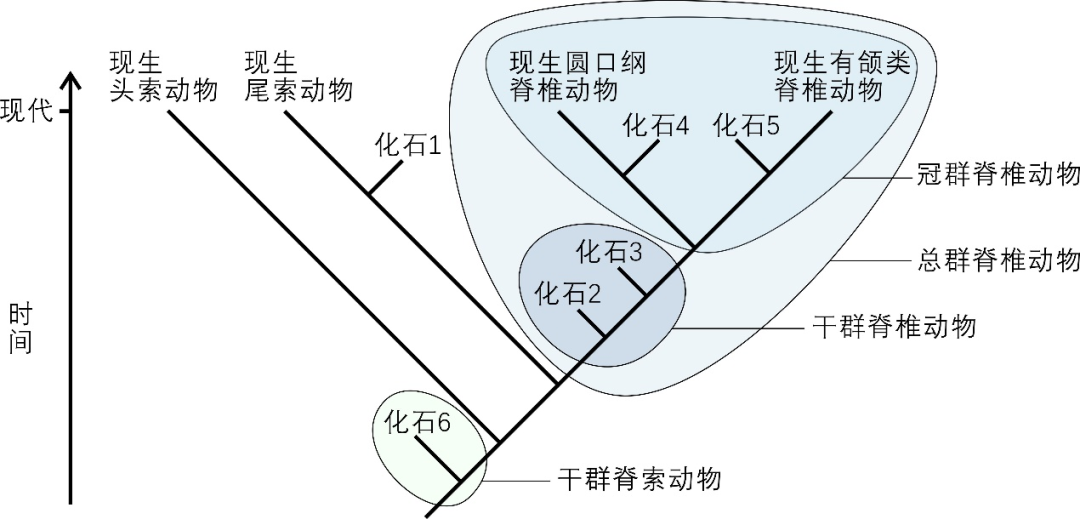
ϵͳ������ܷ�ӳ��������Ⱥ֮�����Ե��ϵ�������ݻ��Ĺ��̣����ִ��ݻ�����ѧ�ձ���õ���Ȼ������ϵ [2]���ڶ���Ҫ������Ⱥ��Դ�������ݻ����о��У�һ����Ҫ�������Ʋ��������ֵ����ͬ���ȣ�Last Common Ancestor��LCA��Most Recent Common Ancestor��MRCA��������Щ��״������Ϊ�˷�������������Ⱥ��Դ�������ݻ����ݻ�����ѧ������˹�Ⱥ����Ⱥ����Ⱥ�ȸ��� [3,4]��
��Ⱥ��crown group��ָijһ������Ⱥ�����������ֵ����ͬ���Ⱥ����Ȳ��������к�������������ִ�ĺ�������ĺ����ͼ1����ijһ��Ⱥ�������ݻ���ϵ����Ĺ�ȺҲ����һ�����ͬ���ȣ�������һ���ȷ������������֧��Ϊ��������Ⱥ�ֱ��Ӧ����Ⱥ��total group������Ⱥ�г�ȥ��Ⱥ����֮�����ڹ�Ⱥ����ġ�����������֣�������Ӧ��Ⱥ��Ӧ�ĸ�Ⱥ��stem group���������һЩ�����ݻ������п���ijij��Ⱥ��������Ⱥͨ��ָijһ��Ⱥ�ĸ�Ⱥ��
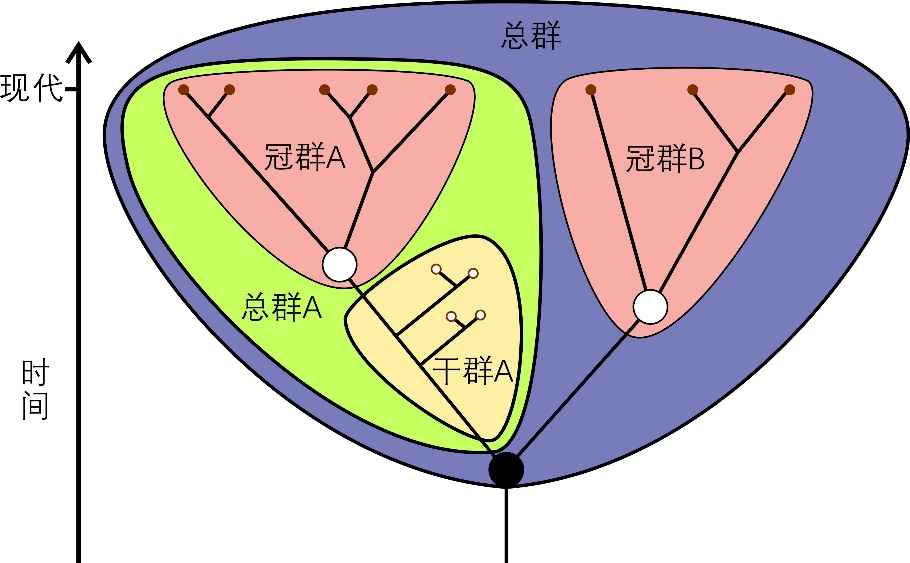
ͼ1 ��Ⱥ����Ⱥ����Ⱥ��ʾ��ͼ��ͼ��չʾA��B������Ⱥ����ȺA��Ӧ�ĸ�Ⱥ����Ⱥ���Լ�A��B������Ⱥ���ɵ���Ⱥ������ʾ������Ⱥ��С���ʾ��ʯ��Ⱥ������ʾ��Ӧ��Ⱥ�����ͬ���ȣ���ڵ��ʾ������Ⱥ�����ͬ���� | ����ά���ٿ��û�Mattbierner��Peter coxhead��https://en.wikipedia.org/wiki/Crown_group��(CC BY-SA 4.0)��
��֤�Ŀ�ѧ��
�������� ���� ���κνϸߵȵ���Ⱥ��Դ����Ӧ������һϵ�нϵ͵���Ⱥ��Ҳ�������ǵ�������Ⱥ���Ǿ����������Ļ����£��Լ��ɻ�����������������ٹ��촴�£���һ��һ���������ġ� ��һ���������������ָ�� ����������ΪijЩ�͵���Ⱥ��Ϊ����ĸߵ���Ⱥ������ijЩ����ͻ������٣��͵ó�����˵ ��ǰ�߾��Ǻ��ߡ�������˵ ���͵ȶ�����Ǹߵȶ����
Ϊ��˵����������ϵ���������� ���о����������ӡ���һ��������Ϊ�������һ֧�ݻ������ж�������ж����һ֧�ݻ������ࡣ�ڶ���������� ��3000������5000������ǰ������ijһ��dz�ԭʼ��С���ӡ� �ݻ�����Գ������Գ�ݻ������ࡣ�����������о��˰��������ݻ�������������������ݻ���������������� �����������ּ����ݻ���ϵ�Dz��Ͻ��ģ�������ϵͳ����������ӳ����ϵͳ������ϵ�����롰���ġ���صİ�������Ϊ�������������ں�ڶ���ϵͳ��������ƫ�����������Ⱥ�ݻ���֧ϵ��ͼ2����Ҳ����˵�������������ݻ�����������ͼ���������������ɼ�������Ͱ�����������ͬ�����ݻ�������������ƣ�����������ƫ������Ĥ�����Ⱥ�ݻ���·�������ݻ������ж�������ࡣͬʱ��������ݻ�Ҳ�����ڴӵͼ������������ݻ�˳��
�������������У��������� ��Ϊ���ܽ��������Ϊ���࣬���ܽ����ӳ�Ϊ���࣬Ҳ���ܽ����������Ϊ������������� ���������������оٵ�����ϵ�� �����ġ� ��ϵ����������Ϊ �������� ��Ϊ�������ϳ漰��ͬ�������������Ĺ����ǣ����ɵ͵ȵ� ����������������ϳ棬���������ţ��ų涯���ŵ������е�ijһ֧�����Ƚ����ɾ��м����� ���͵ȼ������������ͷ�����β��������ټ����������㴴���ͷ�ԣ�������������ճ�Ϊ�������������ݽ�·����ѧ�Ҹ���൱����ˡ��� ��������û���вο����ף������������Ϊ���ֻ����ݽ�·������൱����Ŀ�ѧ����ָ��Щѧ�ߡ����ڹų涯�vetulicolians�������ϳ棨yunnanozoans�����������������������ͼ�����֮���ϵͳ������ϵ���ھ���������µġ��йؼ�������Դ�������У�������Ϊ������� [5�C7]����һ���� �����ġ� �����Բ�������˵�������ϳ��ڲ�ͬ�о��б����ڼ������Ⱥ [8,9]��ͷ������ [10]���������� [11]����ڶ����Ⱥ [12]������ԭʼ������Գƶ��� [13]��
�����ݻ��漰�ڶ���Ⱥ����״�ĸ��ӹ��̣������ôӵͼ������ġ���������������ϵͳ�������������ۺϲ�ͬ������Ⱥ���ڶ���״�������ݣ������ʺϵ��������㷨�������ؽ������ݻ��Ĺ��̡������ġ� ��ϵͳ����������������Ժʿ��2017�귢������ƪ����[14,15]�е������״���ݺ��йط������������ڴ˻����ϳ�ֿ����˵�ǰ�о��л�ʯ����ѧ��������ϵIJ�ȷ���Ժ����飬��ȡ�˱��ص���״���뷽ʽ���� �����ġ� �����ĸ����������õ�ǰ�о��г��õ��Ը���ģ��Ϊ�����ı�Ҷ˹�ƶϷ����ó������ϳ������λ��ͼ2��ʾ���ݻ�λ�á�
��Ȼ������Խ���ѧ��״���ò�ͬ�۵�Ľ��ͺ����ݱ��룬���߲��ò�ͬ��ϵͳ��������������ģ�� [16,17]�����ܻ�ó��� �����ġ� ����в����ϵͳ������ϵ�����DZȽϺ���֤��ͬ�ݻ������г���������
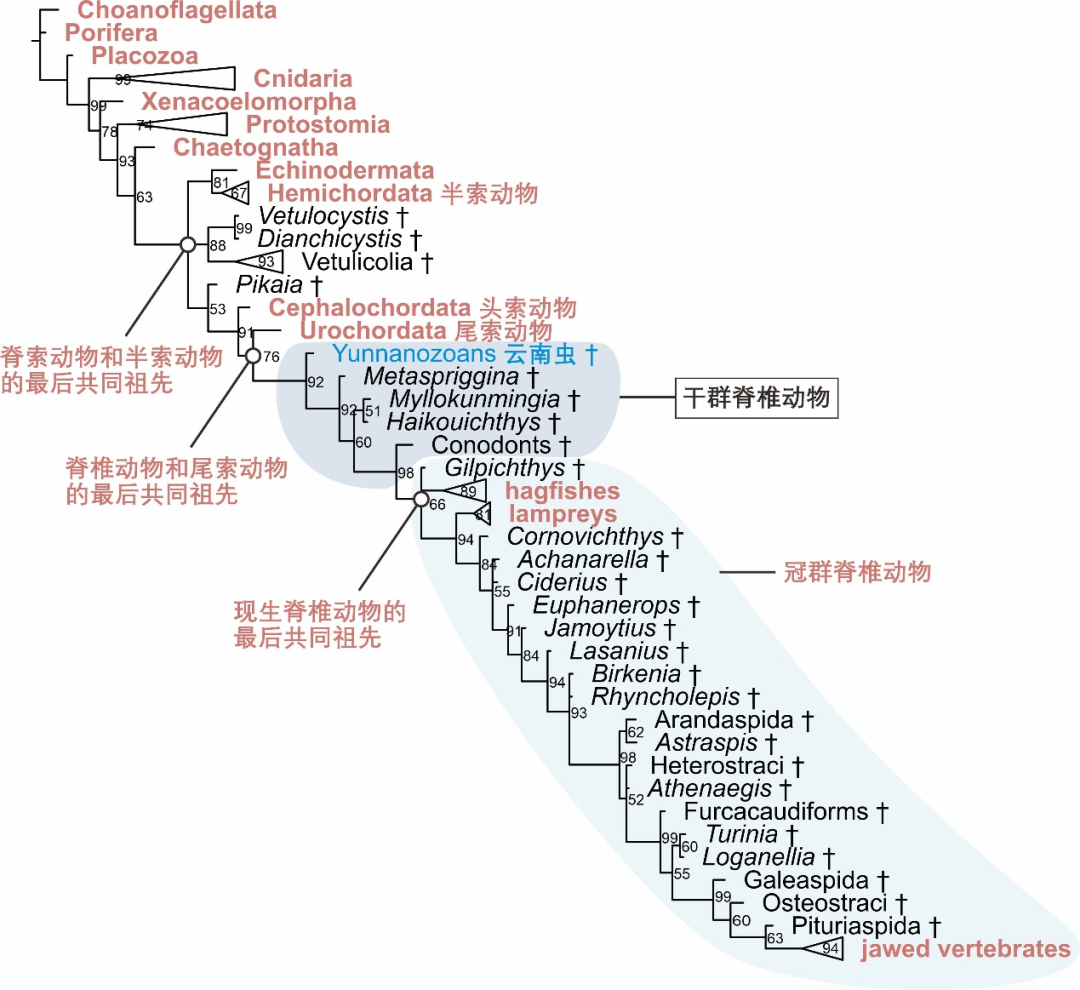
ͼ2 �����ġ� �еõ���ϵͳ��������������Ⱥ��ʾΪ��ɫ�������Ⱥ�ý��꣨†����ʾ��������Ⱥ�ϲ���ʾ���ڵ��Ե����ֱ�ʾ������ʣ��ٷֱȱ�ʾ����
�����������ͷ�����β������ͼ��������ϵͳ�����о��������������β������֮��Ĺ�ϵ�������ͷ���������ڼ���������������������һ֧ [18]����ǰ���ᵽ�Ĺ�Ⱥ����Ⱥ�ĸ���Ӧ�õ���������Դ�������У����Կ���������Ĺ�Ⱥ����������Բ�ڸٺ�����ࡣ��������ĸ�Ⱥ��������ڼ�������Ⱥ֧ϵ�ϡ�λ�ڼ������β���������ͬ���Ⱥ�Ⱥ������֮��������Ѿ�����Ⱥ��ͼ2���������ġ� ͨ��ϵͳ����������ʾ�����ϳ�����ݻ�λ�ô��ڼ�����ĸ�Ⱥ֮�У�������Ŀǰ��֪��������ļ������Ⱥ֧ϵ��ͼ2���������ġ� ��δ���������ͷ�������β�������Ϊ�����
ͼ3 ��������ϵͳ������ʾ��ͼ | ��������[4]����
֤�ݵĿ�ѧ��
�������� ��Ϊ�������ϳ���˾��������⣬���߱�����ؼ������������������û�м�����û�м��ڣ�û�иغ�β��û�м��ǣ�Ҳû��ͷ���ۡ����ԣ����ϳ��뼹�������ϵ����
���ȣ�������Ҫ������Ⱥ����Դ���ݻ��о���������Ⱥӵ�е�һϵ����״�����������ڸ���Ⱥ��Դ�������ݻ�����õ� [19]��ijһ��Ⱥ�ĸ�Ⱥ���֣�����ֻ�������Ⱥ��������״�������������ݻ������еĹ������͡����ϳ治���й�Ⱥ�������һЩ��״������Ҳ�������ϳ����ڸ�Ⱥ�����
��Σ���ʯ�����ǻ��ŵ������������������õ�����������ɻ�ʯ���ᾭ�������ĵ���ʱ����ӵ������ѧ�������仯 [20]������������У��ܶ�������������ʧ���������µ�֤��û�б��۲쵽�����Ի�ʯ���������Զ�������������ˣ���ʯ��ȱʧ����������ȷ�������Ƿ���ڵģ�����������֯���� [5]�����ϳ滯ʯҲ�������Ƶ���������ϳ滯ʯ�������۽ṹ����ͬ�о��Ŷ��в�ͬ�Ľ��͡����磺
1.��������������ļ������ɺ��н�ԭ�������ʺ��ڲ���ϸ����ɵ� [21]����Ȼ�������� [6]�����ǿ�����Ϊ������̼���������ڻ�ʯ�в������� [20]����ʯԲ�ڸټ�����ϣ����ä����Myxinikela siroka��[22,23]���ɴ��ù���˹������Hardistiella Montanensis��[24]��ղ���˫����Pipiscius zangerli��[25,26]������������Ĵ庣���㣨Haikouichthys ercaicunensis���걾 [27,28] ��û�й۲쵽������������Ϊû�й۲쵽�����ͷ�����Щ��ʯ�Ǽ������ǰ��Ҳ��������ϳ漹���Ľ��� [8�C10, 29, 30]����Ȼ�������飬���ǻ����ܼض������ϳ�û�м�����
2.���ڡ������Ƿֶε����ɼ��⣬���������м������ﶼ�е�������ͷ������ͼ�����ӵ����̬��ͬ�ļ��ڣ���β������û�м��� [31]�����ԣ����ܸ���û�м������϶����ϳ治�Ǽ���������ϳ汳���ķֽڱ�ǰ�˽���Ϊ���ڣ�Ҳ�����м�����ά�Ĺ��� [8�C10, 29, 30]�����ܴ������飬���Dz������������еļ���֤�ݡ���ʹ�����µ�֤��֤�������ֽ��DZ�Ƥ�ṹ��Ҳ�����ų����ϳ��ڱ�Ƥ֮�¼����������ڵĿ����ԡ�
3.�غ�β�����ϳ�����ɺ���������Ե�ͻ���� [8,9,32]���ýṹ��ǰ�˽���Ϊ�غ�β [8,9]����������ѧ�߶Ըý������������ [33]��
4.�������Ǹ����ڼ����ϷֽڵĹ����������ǡ����е�Բ�ڸټ������У�ֻ���������������������ä������Eptatretus����β�� [34]��������Ļ�ʯԲ�ڸ�Ѱ������������Tullimonstrum gregarium��[35] �м���������ä������Myxine��[36] ����������Բ�ڸٻ�ʯ��û�з��ּ���������Ϊ��Щ������û�м���˵���Dz����ڼ����
5.ͷ���ۡ�����ͷ�����������ˣ�Lamarck, 1801��[37]�Ķ��壬������ͬ�������ͷ�ࣨCraniota Haeckel, 1866��Craniata Lankester, 1877��[38, 39]����ζ�����Ƕ��й��ʻ������ǵ�ͷ��ͷ������Χ���Ե����Ͱ�Χ��ǻ�������������е�ͷ������Ͱ�������Ҳ��֧����ǻ�������ṹ������������������ɵġ������ġ� �ṩ�����ϳ�����ʲ����ǵ�֤�ݣ�˵�����ϳ������������֧�ŵ���ǻ����ԭʼ�������ݴ�ͳ�ϼ�����Ķ��壬���ϳ��Ѿ������˹ؼ��ļ�����������
������Ҫ������������֯��ɣ�ͬʱҲ�Ǹ�����������֮һ [6]������������ﻯʯ��û�б�����ԡ������۾������������кܶ����۾���ͷ�������β����������ǰ�˶�����С�ĸй����٣��ּ����������������� [40]�����ǣ��༹ܶ����Ҳ���������Ե��۾�����������ֻ�����������������ä�������ǵ��۾�û�о�״��ͺ�ɫ�أ���ȫ��Ƥ������ [41]����ʯ��������˹ä����Tethymyxine tapirostrum��Ҳû�й۲쵽�۾� [42]����ʵ�ϣ�ǰ���������������ϳ�걾�Ͼ����۾����� [8,9,30]��ֻ�DZ����۾��ı걾���٣��Ŵ������顣������ϳ���۾����߸й�������ä������ͷ���������������ԣ������ڻ�ʯ�б�����ҵ��۾��ĺۼ���
���⣬�������� ������ᵽ���������½����ϳ���뼹�������Ҫ�۹���֤���ǣ�������߾������Ƶ� ��ϸ�����ǡ�����������״���ж���ԺͲ�ȷ���ԣ��ܿ��ܹ㷺�������ڶ���������Ⱥ���������ġ� ��û�з���������Ҳ�����ǣ�����Ҳ�ٳ���ͷ������������ǵ����� [1]�������ġ� �ṩ����Ҫ��֤�ݲ������ϳ����ϸ�����ǣ��������ϳ�����˼��������е����������������ǣ���Ҳ�� �����ġ� �ĺ���֤�ݡ�
��ѧ�о���Ҫ����̽��
�ҷdz���л��Ժʿ���Զ� �����ġ� �к�ý�������д��ڵ��������������Ҳ���Զ���Ժʿ�����һЩ���ɱ����˲�ͬ�۵㡣�������������Ŀ�ѧ��һֱ���������еķ�ʽ�����ң�Ҫ�Ͻ��ضԴ���ѧ���⣬���裬С����֤���ҽ������ġ���ѧ�о���Ҫ����̽������Ϊһ֧�������Ŀ����Ŷӣ����dz��Բ��ö�ѧ�ƽ���ķ�����Ϊ��������������ڵ��������⣬�ṩһЩ��̬ѧ�ͳ��ṹ�����֤�ݡ�����һ�����У����ǿ϶������������������IJ��㡣
�����ġ� �Ľ��۽��������ǵĹ۵㣬һ���кܶ�ѧ�߳��в�ͬ�۵㡣ϣ�� �����ġ� ������ש�����Ч��������ͬ������ѧ���������ԵĹ۵㡣����δ�����и����ͬ�й�ע��һ��������������һ���ᷢ�ָ������֤�ݡ���֤��Ҳ��֤�����ǵĽ����Dz�ȷ�ģ������ǻ���ϣ�������Ƿ��ֵ���Щ֤�����м�ֵ�ġ����Ҳ��л��֪ʶ���ӡ��༭���ܸ���������һ�����ᣬ����Ժʿ�����߱��ﲻͬ��ѧ���۵㡣
�ο����ף�
1. Q. Y. Tian, F. C. Zhao, H. Zeng, M. Y. Zhu, B. Y. Jiang, Ultrastructure reveals ancestral vertebrate pharyngeal skeleton in yunnanozoans. Science. 377, 218�C222 (2022). https://doi.org/10.1126/science.abm2708
2. E. O. Wiley, B. S. Lieberman, Phylogenetics: Theory and Practice of Phylogenetic Systematics, Second Edition (John Wiley & Sons, Inc., Hoboken, NJ, USA, 2011). http://doi.wiley.com/10.1002/9781118017883
3. G. E. Budd, S. Jensen, A critical reappraisal of the fossil record of the bilaterian phyla. Biol. Rev. 75, 253�C295 (2000). https://doi.org/10.1111/j.1469-185X.1999.tb00046.x
4. R. P. S. Jefferies, ��The origin of chordates-a methodological essay�� in The Origin of Major Invertebrate Groups, M. R.House, Ed. (Academic Press, London, 1979), pp. 443�C477. https://archive.org/details/originofmajorinv0000unse/page/443
5. P. C. J. Donoghue, M. A. Purnell, Distinguishing heat from light in debate over controversial fossils. BioEssays. 31, 178�C189 (2009). https://doi.org/10.1002/bies.200800128
6. R. S. Sansom, S. E. Gabbott, M. A. Purnell, Non-random decay of chordate characters causes bias in fossil interpretation. Nature. 463, 797�C800 (2010). https://doi.org/10.1038/nature08745
7. P. Janvier, Facts and fancies about early fossil chordates and vertebrates. Nature. 520, 483�C489 (2015). https://doi.org/10.1038/nature14437
8. J. Y. Chen, D. Y. Huang, C. W. Li, An early Cambrian craniate-like chordate. Nature. 402, 518�C522 (1999). https://doi.org/10.1038/990080
9. J. Mallatt, J. Y. Chen, Fossil sister group of craniates: predicted and found. J. Morphol. 258, 1�C31 (2003). https://doi.org/10.1002/jmor.10081
10. J. Y. Chen, J. Dzik, G. D. Edgecombe, L. Ramsköld, G. Q. Zhou, A possible Early Cambrian chordate. Nature. 377, 720�C722 (1995). https://doi.org/10.1038/377720a0
11. D. G. Shu, X. L. Zhang, L. Chen, Reinterpretation of Yunnanozoon as the earliest known hemichordate. Nature. 380, 428�C430 (1996). https://doi.org/10.1038/380428a0
12. D. G. Shu, S. Conway Morris, Z. F. Zhang, J. N. Liu, J. Han, L. Chen, X. L. Zhang, K. Yasui, Y. Li, A new species of yunnanozoan with implications for deuterostome evolution. Science. 299, 1380�C1384 (2003). https://doi.org/10.1126/science.1079846
13. R. A. Dewel, Colonial origin for Eumetazoa: major morphological transitions and the origin of bilaterian complexity. J. Morphol. 243, 35�C74 (2000). https://doi.org/10.1002/(SICI)1097-4687(200001)243:1%3C35::AID-JMOR3%3E3.0.CO;2-%23
14. Q. Ou, J. Han, Z. F. Zhang, D. G. Shu, G. Sun, G. Mayer, Three Cambrian fossils assembled into an extinct body plan of cnidarian affinity. Proc. Natl. Acad. Sci. U.S.A. 114, 8835�C8840 (2017). https://doi.org/10.1073/pnas.1701650114
15. J. Han, S. Conway Morris, Q. Ou, D. G. Shu, H. Huang, Meiofaunal deuterostomes from the basal Cambrian of Shaanxi (China). Nature. 542, 228�C231 (2017). https://doi.org/10.1038/nature21072
16. M. N. Puttick, J. E. O��Reilly, D. E. Pisani, P. C. J. Donoghue, Probabilistic methods outperform parsimony in the phylogenetic analysis of data simulated without a probabilistic model. Palaeontology. 62, 1�C17 (2019). https://doi.org/10.1111/pala.12388
17. P. A. Goloboff, A. Torres, J. S. Arias, Weighted parsimony outperforms other methods of phylogenetic inference under models appropriate for morphology. Cladistics. 34, 407�C437 (2018). https://doi.org/10.1111/cla.12205
18. F. Delsuc, H. Brinkmann, D. Chourrout, H. Philippe, Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature. 439, 965�C968 (2006). https://doi.org/10.1038/nature04336
19. D. H. Erwin, Novelty and innovation in the history of life. Curr. Biol. 25, R930�CR940 (2015). https://doi.org/10.1016/j.cub.2015.08.019
20. L. A. Parry, F. Smithwick, K. K. Nord��n, E. T. Saitta, J. Lozano-Fernandez, A. R. Tanner, J. B. Caron, G. D. Edgecombe, D. E. G. G. Briggs, J. Vinther, Soft-bodied fossils are not simply rotten carcasses - toward a holistic understanding of exceptional fossil preservation. BioEssays. 40, 1700167 (2017). https://doi.org/10.1002/bies.201700167
21. G. Annona, N. D. Holland, S. D��Aniello, Evolution of the notochord. EvoDevo. 6, 1�C13 (2015). https://doi.org/10.1186/s13227-015-0025-3
22. S. E. Gabbott, P. C. J. Donoghue, R. S. Sansom, J. Vinther, A. Dolocan, M. A. Purnell, Pigmented anatomy in Carboniferous cyclostomes and the evolution of the vertebrate eye. Proc. R. Soc. B Biol. Sci. 283, 20161151 (2016). https://doi.org/10.1098/rspb.2016.1151
23. T. Miyashita, A Paleozoic stem hagfish Myxinikela siroka �� revised anatomy and implications for evolution of the living jawless vertebrate lineages. Can. J. Zool. 98, 850�C865 (2020). https://doi.org/10.1139/cjz-2020-0046
24. P. Janvier, R. Lund, Hardistiella Montanensis n. gen. et sp. (Petromyzontida) from the Lower Carboniferous of Montana, with remarks on the affinities of the lampreys. J. Vertebr. Paleontol. 2, 407�C413 (1983). https://doi.org/10.1080/02724634.1983.10011943
25. D. Bardack, E. S. Richardson, New agnathous fishes from the Pennsylvanian of Illinois. Fieldiana Geol. 33, 489�C510 (1977). https://doi.org/10.5962/bhl.title.5167
26. T. Miyashita, R. W. Gess, K. Tietjen, M. I. Coates, Non-ammocoete larvae of Palaeozoic stem lampreys. Nature. 591, 408�C412 (2021). https://doi.org/10.1038/s41586-021-03305-9
27. D. G. Shu, H. L. Luo, S. Conway Morris, X. L. Zhang, S. X. Hu, L. Chen, J. Han, M. Zhu, Y. Li, L. Z. Chen, Lower Cambrian vertebrates from south China. Nature. 402, 42�C46 (1999). https://doi.org/10.1038/46965
28. X. G. Zhang, X. G. Hou, Evidence for a single median fin-fold and tail in the Lower Cambrian vertebrate, Haikouichthys ercaicunensis. J. Evol. Biol. 17, 1162�C1166 (2004). https://doi.org/10.1111/j.1420-9101.2004.00741.x
29. J. Dzik, Yunnanozoon and the ancestry of chordates. Acta Palaeontol. Pol. 40, 341�C360 (1995). https://bibliotekanauki.pl/articles/19946
30. J. Y. Chen, ��The origins and key innovations of vertebrates and arthropods�� in Earth and Life (Springer Netherlands, Dordrecht, 2012), pp. 239�C379. http://link.springer.com/10.1007/978-90-481-3428-1_10
31. T. Onai, The evolutionary origin of chordate segmentation: revisiting the enterocoel theory. Theory Biosci. 137, 1�C16 (2018). https://doi.org/10.1007/s12064-018-0260-y
32. P. Y. Cong, X. G. Hou, R. J. Aldridge, M. A. Purnell, Y. Z. Li, New data on the palaeobiology of the enigmatic yunnanozoans from the Chengjiang Biota, Lower Cambrian, China. Palaeontology. 58, 45�C70 (2015). https://doi.org/10.1111/pala.12117
33. D. G. Shu, S. Conway Morris, Response to comment on ��A new species of yunnanozoan with implications for deuterostome evolution.�� Science. 300, 1372�C1372 (2003). https://doi.org/10.1126/science.1085573
34. K. G. Ota, S. Fujimoto, Y. Oisi, S. Kuratani, Identification of vertebra-like elements and their possible differentiation from sclerotomes in the hagfish. Nat. Commun. 2, 373�C376 (2011). https://doi.org/10.1038/ncomms1355
35. V. E. McCoy, E. E. Saupe, J. C. Lamsdell, L. G. Tarhan, S. McMahon, S. Lidgard, P. Mayer, C. D. Whalen, C. Soriano, L. Finney, S. Vogt, E. G. Clark, R. P. Anderson, H. Petermann, E. R. Locatelli, D. E. G. Briggs, The ��Tully monster�� is a vertebrate. Nature. 532, 496�C499 (2016). https://doi.org/10.1038/nature16992
36. K. G. Ota, Y. Oisi, S. Fujimoto, S. Kuratani, The origin of developmental mechanisms underlying vertebral elements: implications from hagfish evo-devo. Zoology. 117, 77�C80 (2014). https://doi.org/10.1016/j.zool.2013.10.010
37. J.-B. de M. de Lamarck, Syst��me des animaux sans vert��bres ou tableau g��n��ral des classes, des ordres et des genres de ces animaux (L��auteur, 1801). https://doi.org/10.5962/bhl.title.14255
38. E. Haeckel, Generelle Morphologie der Organismen (1866). https://www.degruyter.com/document/doi/10.1515/9783110848281/html?lang=en
39. E. R. Lankester, Memoirs: notes on the embryology and classification of the animal kingdom: comprising a revision of speculations relative to the origin and significance of the germ-layers. J. Cell Sci. s2-17, 399�C454 (1877). https://doi.org/10.1242/jcs.s2-17.68.399
40. S. N. Bradshaw, W. T. Allison, Hagfish to illuminate the developmental and evolutionary origins of the vertebrate retina. Front. Cell Dev. Biol. 10, 1�C20 (2022). https://doi.org/10.3389/fcell.2022.822358
41. E. M. Dong, W. T. Allison, Vertebrate features revealed in the rudimentary eye of the Pacific hagfish (Eptatretus stoutii). Proc. R. Soc. B Biol. Sci. 288, 1�C21 (2021). https://doi.org/10.1098/rspb.2020.2187
42. T. Miyashita, M. I. Coates, R. Farrar, P. Larson, P. L. Manning, R. A. Wogelius, N. P. Edwards, J. Ann��, U. Bergmann, A. R. Palmer, P. J. Currie, Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological�Cmolecular conflict in early vertebrate phylogeny. Proc. Natl. Acad. Sci. 116, 2146�C2151 (2019). https://doi.org/10.1073/pnas.1814794116
��ر��������� | ��¸ɣ����ȳ������ϳ棬�����Dz��Ǽ����

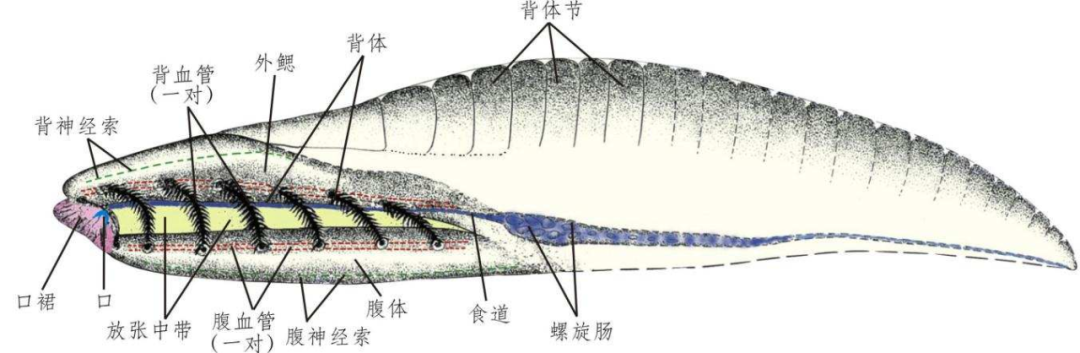
���ϳ渴ԭͼ ���
��֩����eeook.com��������֪ʶ���ӣ��� ��¸ɣ������߰�������7��8�գ�����ѧ����Science����־������һ�����й���ѧԺ�Ͼ����ʹ������о������Ͼ���ѧ������ɵ��о��ɹ�����ͬ���䷢��һλ����ͬ��д�Ķ�������ƪ������Ҫ�о��˾��5.18����ijν�����Ⱥ���ֵ����ϳ���ʹ����еij��ṹ��������Ϊ�����״ϸ�����Ǽ�ԭ��ά����������ϸ�������Ϊ�����������С�ͬʱ�������������µ�ϵͳ���������ó����ۣ���Ϊϵͳ����λ�ó��ڴ�����������ϳ�Ϊ��������ԭʼ����Ⱥ�����·���������ý����д���������Ҳ���������ͬ�е� �����顱�����գ�������ѧ��¸�Ժʿ��֪ʶ���ӡ�Ͷ�壬�ʹ˷������Ŀ�����
����ܸ��˸���ҷ���һ����Ȥ�Ļ��⣬���ڿ�ѧ��Ҳ��һ������Ҫ�Ŀ��⡣֮����˵����Ҫ������Ϊ���漰���������࣬��Ȼ��ֱ�������ִ�������Դ�������������������ļ������������ͥ��Դ������ء�
��Ҷ�֪��������200��ǰ����1822�꣬����������������������ѧ�������ˣ�Jean-Baptiste Lamarck�����ͽ���������绮��Ϊ����������������֡�����ʱ����������������δ����������ݻ���������һֱ�ܵ�ѧ���ǵ��ر��ע��

ͼ1�����۵�����������������ѧ��������
�ڽ���������ܹ�ע����Ŀ������ص����ۣ������ϳ澿���Dz��Ǽ������
����̸������ⲻ����Ѫ���������³�������Ϊǰ�����ҹ��м���ѧ�ߣ���������һ��������־Science������ѧ�����Ϸ�����һƪ���ģ������ϳ����������̬ѧ���о����ó���һ����Ϊ�䶯�Ľ��ۡ��� �����ϳ���ԭʼ�ļ������

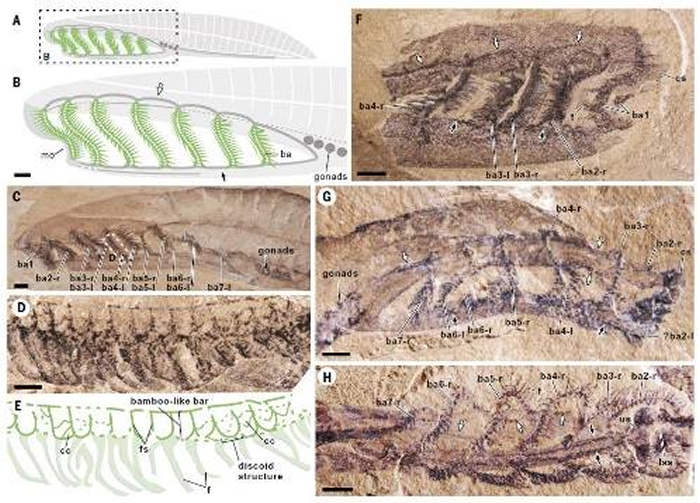
ͼ2 2022��7��15�ճ���ġ���ѧ���������˹������ϳ���о�����
�����о���ֵ�ù����ģ�����Ҳ����һ���Ŀ�ѧ�ο���ֵ�����۵�˵������о�����̫���죬֤�ݺͽ��۶����ڱȽϴ�����⡣�ر��ǣ��������������ڸ���ý�峴�÷з������ѧ�������������Ե����б�Ҫ���Թ������塣
��֮����˵����о������⣬��Ҫ�ǻ������¼������棺
��һ����̸�����ϳ����Щ�۽ṹ���� ������ԡ���Ҳ���Ǿ��кܴ�� ����ȷ���ԡ��������ж��ֽ��͡�����ж��ֽ��ͣ��Ϳ��ܵó� �����ֽ��ۡ���
�ڶ������µ��������⡣��ѧ�������ǽ�������ѧ�Ͼ��������� �������� �������˽���һ��dz���Ҫ��
���������� ��֤�ݡ�����ʵ֤��������Ҫ�����ݲ��ɿ���֤�ݣ��������Ȼ���ɿ���
�ҹ������ϳ��������о���ѧ���Ŷ���Ҫ���ĸ������˸����Ŷ��⣬���������ŶӶ���Ϊ �����ϳ治�Ǽ�������ұ���Ҳ��Ϊ���ó� �����ϳ���ԭʼ����� ������ۣ��൱�����ס�
���죬����ҪΧ���ſ�ѧ���Ϳ�ѧ֤���������������ۡ�
��֤�����Ͽ�ѧ��
������̸��һ�����⣬������������Ļ�������ijһ����Ⱥ��ζ�λ�Ļ������
ʲô�Ǽ�������Դ�ݻ��Ļ������أ������Ƚ��ܽ�������ѧ�е�һ������ԭ�������ǿ�ѧ�繫�ϣ��κνϸߵȵ���Ⱥ��Դ����Ӧ������һϵ�нϵ͵���Ⱥ��Ҳ�������ǵ�������Ⱥ���Ǿ����������Ļ����£��Լ��ɻ�����������������ٹ��촴�£���һ��һ���������ġ�����˵���ߵ���Ⱥ���ɵ͵���Ⱥ���������ģ�������һ�����ͣ�������������һ�콨�ɵġ�
��Ȼ�����Ǿ�������ΪijЩ�͵���Ⱥ��Ϊ����ĸߵ���Ⱥ������ijЩ����ͻ������٣��͵ó�����˵��ǰ�߾��Ǻ��ߡ�������˵���͵ȶ�����Ǹߵȶ��������������Ȼ�Dz������ġ�
�Ҿټ������Ӱɡ���һ�����ӣ�����������ݻ������࣬���ϵ��ݻ�·���ǣ��Ƚ����������࣬Ȼ������������ࡣ������ոմ�ˮ�����ϰ������Ĺ���dz�ԭʼ����������������·��Ȼ�������Ĥ�ѣ������ݽ��������࣬��һ������½�ء��ٺ���������ijһ֧��ǰ֫�ݽ��ɳ���һ�����ë�ĸ��������ܹ����ɷ����ˣ��Ž����������ࡣ��ʱ�������ܲ���˵�������࣬����˵���ܰɣ������������أ��������һ����������˵���ܻ�����Ц������ȻΥ���˿�ѧ����һ��ij�ʶ��
���پٸ����ӣ������������ȣ�����˵3000������5000������ǰ������ijһ��dz�ԭʼ��С�������ݻ����ġ���䵱Ȼ�������˹�Գ�������Ϸ���Գ�Σ����ŵ������ࡢ�����˿ơ������������֡�����������̴�Ҷ�����Ϥ����ô�������ܲ���˵�����Ӿ��������ء�����Ȼ���ܣ���Ϊ��Υ���˿�ѧ�������Ǹ����ԵĴ���Ҳ��ԭ���ԵĴ���
���پٵ��������ӣ����漰���������Ŀ�������ϳ漰��ͬ�������������Ĺ����ǣ����ɵ͵ȵġ���������������ϳ棬���������ţ��ų涯���ŵ������е�ijһ֧�����Ƚ����ɾ��м����� ���͵ȼ������������ͷ�����β��������ټ����������㴴���ͷ�ԣ�������������ճ�Ϊ�������������ݽ�·����ѧ�Ҹ���൱����ˡ�
���ϳ滯ʯ��ų涯���Ż�ʯ�Ĺ���ʮ�ֽӽ��������������İ��������Ŷ��dz�ԭʼ�����ơ��Ұ��������幹������ �����ޡ�����ȱʧ������������������ڣ��غ�β�����ǣ���ͷ�ԡ������Ƚ����� ���͵ȼ����������ͷ�������IJ����β�����ﺣ�ʣ���������ݽ��� ���ߵȼ���������� ���������������������� ����������������Ϊ �����С� �������������������м������������μ��ڣ��иغ�β������Ҫ�����м��ǡ���ͷ�����ˡ�
�������ݽ�·���£���ǰ����������һ���������ܲ���˵�����ϳ���Ǽ������أ���Ȼ���ܣ���ͬ���Ǹ����Դ���
ȱ���ɿ��Ŀ�ѧ֤��
�о���������Դ��������·������һ�����ۣ��ڶ����Ǻ�ۡ�
���ϣ����ǿ����Ļ������һ��һ�����£�����ؼ�����ͬԴ����Ĵ��¡��ں���ϣ����ǿ��������������£������������ٴ��µ�֤���ˡ�
��̸̸��������Ϣ�����������һ�ֵ��ػ���Ҳ�з����ܵ��ػ�������������ж������̥������ϵͳ�����м�����������õ�һ���������˰����п��Ʒ����� ���ܿ��ء�������ѧ�����ֽ�����ͬԴ���ͻ��������� ��ͬԴ�������Ӣ�Ľ� Hox gene cluster�����ֶ������ֻ����һ��ͬԴ�������һ�������������������ʮ�����������������ź͵͵ȼ��������ֻ��һ����������˼���ǣ�������������ʱ������ ��ͬԴ����� �����˾�� ����������ͬԴ����㷢չ���˶��������Ӳ�����������������������ĸ��������ǽ���������� ��ͬԴ����������
��� ������� �dz��˲�������ֻ��һ���������������࣬Ҫ���öࡢ���ӵöࡣ����ͬԴ����Ķ���������˼�������Դ���ڶ��������Ǿ��γ��˼��ǣ��γ���ͷ���Ժ��۾�����һ����Ϊ�ؼ������ڻ����������ѱ���Ϊ��ʯ��Ŀǰ��������Ȼ���˽�������ǰ��ʯ�Ļ����ݻ������ֻ�����ݻ�ʯ�����������С�
�Ӻ���ϣ������ٿ�������������������������һ��һ���������������µġ�
�����������������ǽ磬�����������ǽ磨�����ڸغ�һ��ˮĸ��ɺ���ȵ͵���Ⱥ����ԭ�ڶ����ǽ磨����20������࣬���֫�����ţ����嶯���š����㶯���ŵȵȣ��ͺ�ڶ����ǽ磨���������������ڵļ�������Ա��ܹ�ע����
���棬���Ǽ��п�һ����ڶ����ǽ��ְ������������أ�������������������ţ��ӵ͵ȵ��ߵȣ��йų涯���š����������š���Ƥ�����š�ͷ�����β������ͼ�������Ƕ�֪������������һ�콨�ɵġ�λ�ں�ڶ����ǽ�������˵��Ǽ�������������ش�����࣬�����پ����������ش�Ĵ��£�����������������
��һ�����£������ѵij��֡�����ĺ�ڶ�������ȵ�������û�����ѵģ�������ԭʼ�ĺ�ڶ�����Ⱥ�Ͳ��������ˣ�����һ����Ҫ���½ڵ㡣���ǣ��ں�ڶ����ǽ��У��ոմ������ѵ���Щ�������࣬���ǵ���������Ȼ�� �����ޡ������������������μ��ڣ��غ�β����������ͷ���ۡ�
�ڶ������£����� �������ࡱ �ݻ��� ���͵ȼ����ࡱ�����DZ㿪ʼ���ּ��������ˣ����ǰ����������IJ��㣬�Լ�����͵Ļ�������ʯ�������������ڵ��IJ���dz����ơ���δ��£������˼̳����ȵ�����֮�⣬���������������ٹ���Ĵ��£��������˼����������μ��ڡ��غ�β��
���������£�Ҳ�����һ������ʱ�����γ��˸ߵȼ������Ҳ���Ǽ��������ͻ�ʯ�а���������֪��������Ŀ���� �����µ�һ�㡱����������˼̳������ᵽ���������ĸ����ٴ���֮�⣬������������ؼ������ٴ��£���ͷ���ۺͼ��ǡ�
��֮������ԭʼ�ĺ�ڶ�����Ⱥ��������������������������¼������Ҽ�ס��
������ѧ�о��顢���ϴ�ѧ�о��鼰��Ϫʦ��ѧԺ�ij°����о��飬һ�������Ŷӣ������ϳ泤���о��Ľ���������ƣ�����Ϊ���ϳ���˾��������⣬���߱�����ؼ������������������û�м�����û�м��ڣ�û�иغ�β��û�м��ǣ�Ҳû��ͷ���ۡ����ԣ����ϳ��뼹�������ϵ��
������ѧ�о���ɼ��������ϳ��ࣨ�������ϳ����ͺ��ڳ������걾��Ŀ��8000��飬�����dz��Ӵ��ұ��������ã������Ƕ������ǰ���������Ѳ��֣���Ϣ����÷dz��á����������о��Ŷ�ӵ�е����ϳ�걾ֻ�м�ʮ�������ٸ���������ѧ�о��ŶӶ���8000���걾��������ϸ���죬��ȡ����ȷ������ѧ��Ϣ�������������ϴ�ѧ�ͳ°����ŶӵĽ��۱ȽϽӽ�����֤ʵ���ϳ��� �����ޡ������������ڣ��غ�β��������ͷ���ۡ���Щ֤��������Ҫ��

ͼ3 ���ϳ滯ʯͼ����ԭͼ����û�п��ŵļ������죬û�������μ��ڣ�û�иغ�β��û�м�����û��ͷ���ۡ�
���⣬�ں�ڶ����ǽ��У������ϳ�����ȼ��ಢ�С���̬�����Ƶ���һ���͵���Ⱥ�ǹų涯���š�����������������ѣ�ʵ���˺�������������Ȼ�� �����ޡ������������ڣ��غ�β��������ͷ���ۡ�
���������� ��������� ������ ���͵ȼ���������������ͷ������������˼��������ںغ�β������Ȼ����û��ͷ���ۣ�û�м��������
��ͷ�������ಢ�е�һ��͵ȼ��������β��������ǵij����������Թ۲쵽 �����С��������ѣ��м������м��ڣ��иغ�β������û��ͷ���ۺͼ����죬��������ͷ�������IJ���һ������Ȼ���� ���������δ�ܽ��뼹������ͥ��
��������������һ���Σ�������ͳ����� �����С� ���١�����˵���������̳������ȵ����ѡ����������ڡ��غ�β������Ҫ���ǣ��´�����ͷ���ۺͼ��ǡ�
�����������ֳɶ��е��������֣�ǰ�沿����ͷ���ۣ��м䲿�ֵ������м�֧�ţ����������ͼ��ڰ��������沿�֣����ĸ��Ų��������ĩ�ˣ���������β����һ�ξ���ĵط������ź����и�β�ͣ��иغ�β������������һ�ද����ˮ���˶�����Ҫ�������٣���������Էdz���Ҫ��
�������㵮���� �����µ�һ�㡱����������Ŀ��Ŀǰ������������㡢��������ӽ��㣩���Ӵ˵����ϾͿ�ʼ�����������ˣ����������������ࡢ�����ࡢ������ΰ�����ĵ�һ����

ͼ4 ����һ�㡱 ������Ŀ�Ļ�ʯ����ԭͼ�����������п��ŵļ������졢�����μ��ڡ��غ�β���������˼��ǣ�ͷ���ۡ�
���潲���ǵ�һ�㡣�ڴ˺�����1000����ļ��ô���˹ҳ���У�����ǰҲ�����˼�������Ļ�������ѧ��������һ��dz��ӽ����� ����˹�������㡱����Ϊ����ʱ�����ҹ��ĵ�һ���������������� ���ڶ��㡱 �ɡ�
�����ں�ڶ����ǽ�Ļ�ʯ��¼����������Դ���������������ش�Ĵ��£�Ҳ������ͼ��ʾ������������
ͼ5 ������ѧ�Ŷӷ�����Ӣ��Nature������Ȼ������־�ϵ����ں�ڶ�����ϵ�ݻ�ͼ������ʾ�����ѡ������������μ��ڡ��غ�β�����ǡ�ͷ���۵����ٹ������ݽ����̡�
��ͼ�Ͽ��Կ��������ϳ���ų涯����һ������ʾ��ԭʼ�ԡ��������������δ��º͵���������ԭʼ�������������࣬���״��˼��Ǻ�ͷ���ۡ�
�Ҿ�����һ���������ٵ����Ӱɣ�������۾������Ǻ�ڶ����ǽ��м��������е�һ����Ҫ���죺��������ѧ�����500��������㡢������ı걾�У���һ�����ϵı걾�����˺ܺõ��۾�����˵���۾����죬�ڳν�����Ⱥ���Ǻ����ױ���Ϊ��ʯ�ġ�
���⣬�ڳν�����Ⱥ�У���֫������۾���ʯ��¼Ҳ�൱�ࡣ���ǣ�����˵���ϳ�������۾��Ļ����ڳν�����Ⱥ��ͬ���Ļ�ʯ�γɺͱ��������£���Ӧ���кܴ�ĸ��ʱ�����Ϊ��ʯ��
��ô�����ϳ滯ʯ�е�����û���۾���ʯ�أ�û�У���һ���걾��û���ҵ���
������ѧһ���ɼ���8000����걾�����Ҷ�����������λ�õ��Ǹ���λ��Ȼ��������Щ�걾��û��һ����ʾ�����۵���˿������
�۾�����������Ҫ�����Ǽ������һ���ؼ���ǡ���Ϊ�۾��ڽ���ѧ�����Ե����ӣ�Ҳ����˵�����Ծ�һ�����۾������м���������ǵ͵ȡ��ߵȣ������۾������Ĵ���֤�������ԡ�
��ô����Ȼ���ϳ�û���۾�����˵����һ��û���ԡ�û���ԣ�֤������ͷ��������ȫû������ͷ�ġ�
Ŀǰ����δ���˷��֣������ϴ������κ�û��ͷ���۵ļ������ʵ���ϣ���Ѩ�С����µ���ȫ�ڰ��ط��� ��Ϲ�ӡ� �������Ȼ���۾����죬ֻ��û���������ܰ��ˡ���
��ô����һ��ʣ�Science�������ߵ�һλǰ����ʦ������η��ģ���θ����ϳ渴ԭͼ�� ��װ�䡱 �˲�ͬ���۾����� �����۾�����Ҳ�� ��С�۾����������Ǽ٣�
������ 20����ǰ�������������ϳ��� ���۾���������������֤��֤α�ˡ�������Щ��ν�� ���۾�����ʵ���ϸ��������ڣ�����������ã������ν���۾� �����֡� �ͱ�ѧ�������ˡ����ڣ�����Ҳ���ϣ����ϳ��ȷû���۾����졣
���Dz���Ҫ�ʣ����ϳ��Ȼû���۾��������ܾ��д��������û���۾��ʹ��ԣ����ǿ����Ǽ�������
���ڣ����ۺ������������dz棬������㡱�����ϳ����������е� ���桱��������Ŀ�Ǽ������еġ���һ�㡱��
���ݽ��ȼ��ϣ����ϳ��뼹���ﻹ�൱ңԶ�������ߺܳ���һ��·�����ܽ��뵽��������ͥ��ʵ���ϣ����ϳ桢�ų涯���ͷ�����β�����ﶼ���������
ͷ���ࡢβ�����ǵ͵ȼ����࣬ѧ������Ϊ������ ����ͷ�ࡱ������ ������� ���롣��Ȼ������Щ ����ͷ�ࡱ ���͵ȵĹų桢����������ϳ棬���ɸ��� ������� �ˡ���Ȼ���ӽ����Ͽ������ϳ��п����Ǽ�����ʼ��ĸ������ȵ�ijһ����֧�������������ֱϵ���ȡ�
һ���Ա�֮�����ϳ�ֻ�� ���桱������ ���������������Ŀ�� ���㡱����Щ �����µ�һ�㡱 ���������� ��ԭʼ���������Ϊ����������ϳ棬����Ϊ������ʼ���������Ƚϣ����������蹵�����ɻ�Ϊһ̸��
��Ȼ��Science�ϵ���ƪ�����ڿ�ѧ�ϻ�����������������⣬���ڱ���ƪ������������
��ѧ�о���Ҫ������ʵ
����20����ǰ�������ҹ��Ĺ�����ѧ�ߺͼ����Թ����ﻯʯһ֪���������ˣ���ͬ�ó� �����ϳ�����Ǽ���� �Ľ��ۡ�����������ס�������ͻ�ʯ��ʵ֤�ݵļ��飬�ܿ�ͱ����ʺ���ѧ�������ˡ�
���ڣ� ��Science����ƪ�����У����ý����������������������㷺��ע�������в������������Ͳ�ʵ��֤�ݣ������˴���Ľ��ۣ�����ѧ��ʹ��ڡ�
��ѧ��һ����ʥ����ҵ����Ҫ���Ͻ��Ŀ�ѧ������ʵ�����ǵĿ�ѧ̬�ȣ�ϣ����ѧ���ܴ�����¼�������Ϊ�䡣
��ӭ��Ҽ��� ���ټ���������лл������������¸�������������վ��Ƶ��Ҫ���ݸ�д���ɣ�
���ߺ��
ǰ���õ�һ�죬��ͻȻ�ڹ��ڸ��� ��Ȩ��ý�塱�� ��ͬһ�� ���䶯����ġ� ���ţ�˵����ѧ����־��һƪ���ģ�֤ʵ �����ϳ���ԭʼ�������ƾ�ҹ�ȥ30�����Գν�����Ⱥ�е� �����ѻ�ʯ�� ���ϳ���о����������ٴα� ������Ϊ�� ������ ����ߵȡ� ������Ⱥ������ҵ�ȷ�����ˡ��ҵ�һЩͬ�н�����Ӧ�ü�ʱΪ��ѧ����������һ�����ĵĿ����ŶӰ����Ҳִ���һ��24���ӵ���Ƶ����7��23�շ���������������վ�ϣ��ܵ��������������ǵĹ�ע�������������־����������Ƶ����Ҫ���ݣ��������Թ�ѧ�����ۣ�����ѧ���Ƿǣ��ٽ���ѧ������չ��
���ϳ���������30�꽻��� �������ѡ���1991�꣬�й����������ѧ�����ȱ�����5.2����ǰ�ν�����Ⱥ������ϳ棬��������������ѧ����λ�á����꣬�ұ�������˵Ķ�������ס�ˣ��Ӵ˽��²���֮Ե������ͬ���ǿ�ʼ�㷺�ռ�������Ϣ��ϣ���ܸ���� ������λ�ò����� �����˺� ���Ҹ��ҡ���
1995�꣬���Ͼ� �����ʺ����ѧ�����ۻᡱ �ϣ����ύ��5ƪ����ժҪ�����ڻ�������ͷ����3�Σ�����һ���ǽ� �����ϳ��Ǻ�ڶ����ǽ�ij�Ա���������й�������ѧ���״ι����ἰ ����ڶ�� ���İ�����ʡ��Ӵˣ��ҿ�ʼ�������Ĵӱ�ʶ���ϳ� ��®ɽ����Ŀ�� ��Ѱ�� �������������ԭʼ����� ��̽�����̡�
1996�괺�������ڡ���Ȼ����־�Ͻ����ϳ����Ϊ5.2����ǰ�� ���ư�������������Ų��ã�����Ӣ�����Ŵ�ѧ�Ŀ�άĪ��˹���ں��������ڡ���Ȼ���ϱ����������ͷ�����ﻪ��������Щ���ְ�ʾ�ң�����ϵļ����ﻯʯ�ܿ��������Dz�Զ�ˡ���Ȼ���������ನ�ۺ�ϲ�ã�1999����һ���������һ��������Ϊ����������ϼ������ڡ���Ȼ��������������ѧԺԺʿ��ά����ͬ�ڿ������� ����ס��һ�㡱 Ϊ��ߵ�����������֡��������ļ��꣬���Ǽ��Ұ��������˵زɼ������ٿ�������ĺ����㻯ʯ�걾�������˶��Ľ���ѧ��Ϣ������2003���ٴ��ڡ���Ȼ����������档ͬ�꣬���ǻ������˵����� ����һ�㡱 �ӽ��㣬�������� ��������Ŀ����
�ڴ��ڼ䣬���Dz����ڡ���Ȼ���ϱ����˴ӵ͵� ��������� �����������ݽ��� ���ߵȼ��������������������е�һ�� ���͵ȼ������ β�����ﳤ�����ʣ����һ��ڡ���Ȼ���ϴ�����һ���µĶ������ࡰ�ų涯���š�����Ϊ��ڶ����ǽ�ײ���ԭʼ��Ⱥ�������ϳ�Ͱ��������Ŵ���λ��ͬһ�����ȼ����ڷ��������ں�ڶ����ǽ�����6����Ⱥ֮��2004�꣬�����ڡ���Ȼ����������ִ�����ѧ��֪����һ�µ����ں�ڶ����ǽ���ϵ�ݻ�ͼ������ڶ��������ס����У�1999�귢�ֵ� ����һ�㡱 ��2001�괴���Ĺų涯��������ɹ��ֱ���ѡ�뵱��� ���й�ʮ��Ƽ���չ��������Ҳ˳�����µؼ����˶���Ľ̿��顣2008�꣬���ڳν�����Ⱥ������ ����һ�������� ����֪����ϰ��Ͽ�������Ⱥ��С������Ⱥ�Ȼ�ʯ����Ĵ�������Ϣ����д���ģ���ʽ����� ����Ļʽ����ʹ��� ��˵��
Ȼ�������ڼ�ѧ������Ȼ����һ���������⣬�������ȷ���� �����Ѷ�� ���ϳ������ѧ��λ����1990�����һ�����������о�С�������Ϊ�����ϳ��ǵ͵ȼ�����������ֽ�������Ϊ������� ����ͷ�ࡱ��2003�괺�������Ŷ��ڡ���ѧ����־�ģ����������ϳ����һ�����֣�����������ѧ�ɼ�����ǧö���ϳ���걾������ȫ�濼�죬ȷ֤���ǼȲ��߱��͵ȼ�������ļ��������ںغ�β����û�м�����ר�е�ͷ���ۺͼ��ǵļ����ˣ�����ȫ��λ���������ϳ��������ѧ��λ����Ϊ��Ӧ���Ǻ�ڶ����ǽ��е�ԭʼ��Ⱥ����͵ȼ�������ߵȼ����������������أ�����Ǽ�������İ��������źų涯����������˺����ϳ���� ��������˵�� �ܿ챻ѧ��������
����ѧ����η����������½����ϳ���뼹�������Ҫ�۹���֤���ǣ�������߾������Ƶ� ��ϸ�����ǡ�����ʵ������ȥ��Զ������Ҫ���ǣ�����״���ж���ԺͲ�ȷ���ԣ��ܿ��ܹ㷺�������ڶ���������Ⱥ�����֫���ɺ���ࡢ���Ͽ�������Ⱥ����ɽ����Ⱥ�ȣ����Ȼ��������۵IJ�ȷ���ԣ����Բ��š�
��κ�Ǽ���֮��ص���Ƶ���ǻ����ҵ��о��Ŷ���1996�����������ڡ���Ȼ���롶��ѧ����־��10��ƪ����Ҫ����ۺ�����������֮������ӭ����ָ����
��֪ʶ���ӡ�������ԣ����ֹ�ƽ���������Ե�ѧ��������Ҳ��������Խ��Խ��������������ģ�editor@zhishifenzi.com